2018年1月2日,硕士研究生李倩和巩凤芹同学在组会上做了文献报告,具体内容如下:
一、报告人:李倩 题 目:Structural insights into marine carbohydrate degradation by family GH16 kappa-carrageenases
报告内容:本文作者来自法国,于2017年10月发表在Journal of Biological Chemistry。卡拉胶是存在于红藻细胞壁中的硫酸化的α-1,3-β-1,4-半乳糖,而硫酸化的碳水化合物的降解模式尚未被广泛研究。本文主要通过晶体结构解析对来自两株亲缘关系较远的菌株(Zobellia galactanivorans和Pseudoalteromonas carrageenovora)的GH16κ-卡拉胶酶的结构及理化性质的比对来探究卡拉胶酶的结构及功能特点。
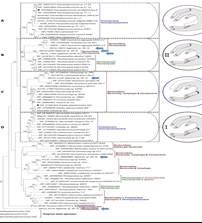
1. 本文首先将从GeneBank 上查找到的76种κ-卡拉胶酶(包括推定的κ-卡拉胶酶)进行多序列比对,构建系统进化 树,根据Bootstrap值选定A、B、C和D四个区域(如下图示)。阐明了四个区域的主要区别。本文的研究对象 Z.galactanivorans和P. carrageenovora 分别来自Clade C和Clade A。

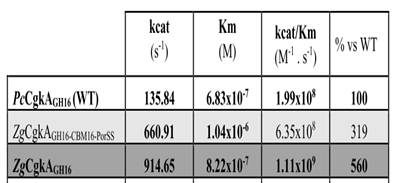
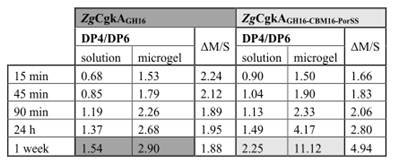
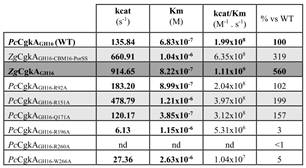
2.克隆表达得到上述两株菌中的酶ZgCgkA、ZgCgkAGH16和PcCgkA。对比了这些酶在卡拉胶稀释液和卡拉胶微凝胶中的催化效率。如下表示:发现ZgCgkAGH16的催化效率是ZgCgkA和PcCgkA的3倍和5倍。CBM16结构域的缺失使反应效率提升了40%。
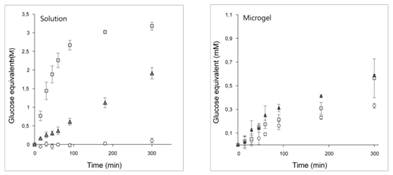
3.作者进一步通过HPAEC对ZgCgkA、ZgCgkAGH16的终产物进行分析,发现两者酶解终产物都是四糖(DP4)和六糖(DP6),ZgCgkA产生四糖量更高。且在微凝胶中,四糖的比例更高。说明CBM16的存在更有助于四糖的生成。如下图

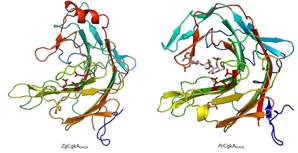
4.构建了失活突变体以获得稳定的酶底物复合物,进而得到晶体,以已经结构解析的PcCgkA作为模型,进行分子置换,得到ZgCgkAGH16的三维结构。并将两者进行比对,如下图:
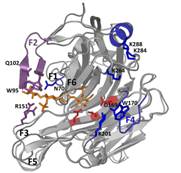
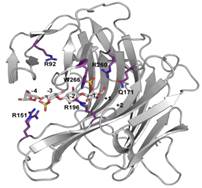
5.为了探究与底物作用的关键氨基酸位点,对PcCgkA进行定点突变,并对突变后的酶进行活性检测,如下所示:
二、报告人:巩凤芹 报告题目:Structuraland Computational Insight into the Catalytic Mechanism of Limonene EpoxideHydrolase Mutants in Stereoselective Transformations
报告内容:
(1)经过定向进化的柠檬烯环氧化物水解酶(limonene epoxide hydrolase,LEH),能够选择性催化环戊烯氧化物或环己烯氧化物,使它们发生不对称水解反应,分别产生(R,R)-和(S,S)-构型的手性二醇。研究人员利用定向进化技术,结合突变体晶体结构分析和理论计算,解析了这种水解酶突变体的底物选择的特异性与立体选择性不对称催化机制。
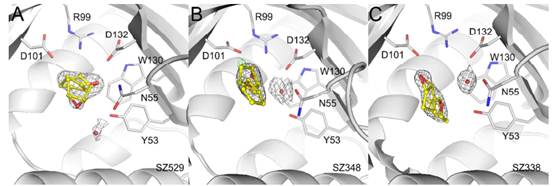
(2)在定向进化过程中,作者发现针对环己烯氧化物筛选出来的突变体,丧失了对环戊烯氧化物的催化能力,而有些突变体则保留了对两者的同时催化性能。通过突变体晶体结构与底物复合体解析,发现在对环戊烯氧化物丧失催化能力的突变体的底物结合口袋中,环戊烯氧化物存在多重构象,不利于催化的发生,MD计算分析发现底物的结合口袋体积发生明显变化,进一步支持了这一结论。一个相对紧凑的结合口袋可以稳定较小的底物和较大的底物,相比之下,更松弛的结合口袋不利于较小底物形成有利的亲和攻击构象,从而导致LEH突变体无法催化较小的底物,而在环氧环己烯的立体反应中,反应构象很容易便能形成。
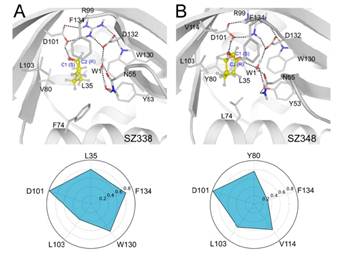
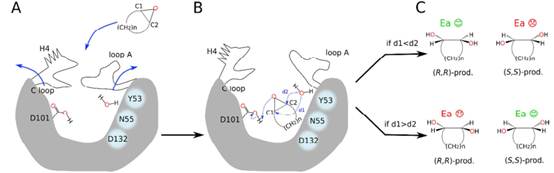
(3)利用立体探针的概念,结合量子力学(QM)和分子动力学(MD)的计算手段进一步阐释了突变体的立体选择性催化机制,当底物接近LEH时,可诱发H4、C环、环A发生构象变化,使底物结合口袋打开,有利于底物进入催化活性中心形成特定几何构型,降低催化能垒。随后,一个已经活化的高度保守的水分子,即与Y53、 N55、D132以氢键结合,通过两种不同的方式进行亲核攻击:要么攻击环氧基的C1基团,要么攻击C2基团。如果水分子距离C1 (d1)基团更近,就有利于(R,R)-构型的手性二醇形成,相反则形成(S,S)-构型的产物。
(4)通过交互指纹(interactionfingerprint,IFP)分析了突变体结合口袋关键氨基酸残基与底物的相互作用强度,它们在维持特定底物构象的形成方面起到至关重要的作用,从而使底物构象与特定的产物相对应。