2017年8月21日,硕士研究生鞠酒和郑远迪同学在组会上做了文献报告,具体内容如下:
报告人:郑远迪报告题目:Highly regio- and enantioselective multiple oxy- and amino-functionalizations of alkenes by modular cascade biocatalysis
报告内容:烯烃易于获得和优异的化学合成原料。烯烃的不对称官能化在药物制造中的对映体化学品的合成中是非常重要的。我们最近开发的烯烃不对称反式二羟基化的环氧化-水解级联是烯烃不对称官能化的唯一已知的生物催化级联。到目前为止,大多数报道的级联生物催化只能进行两到三个相对简单的酶反应。设计包含四个以上酶反应的有效级联系统是非常有挑战性的。末端烯烃分别制备手性α-羟基酸,1,2-氨基醇和α-氨基酸的单釜区域立体选择性多重氧基和氨基官能化被设计为目标反应。
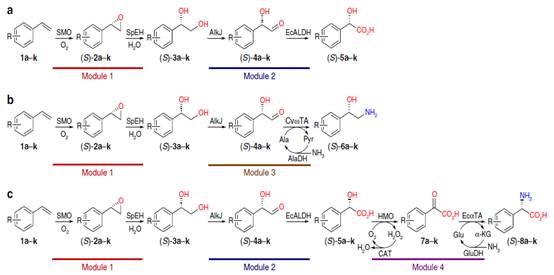
a.分别将含有多个碱性酶模块的末端烯烃一单胺转化为含有多个碱性酶模块的大肠杆菌细胞的手性α-羟基酸,1,2-氨基醇和α-氨基酸。
b.四种一般基本酶模块及其级联生物转化
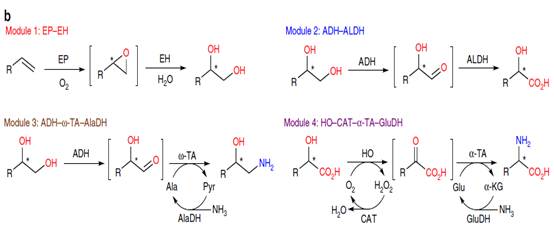
c.通过模块化级联生物催化方法进行对映选择性和对映选择性的苯乙烯氧化和氨基官能化
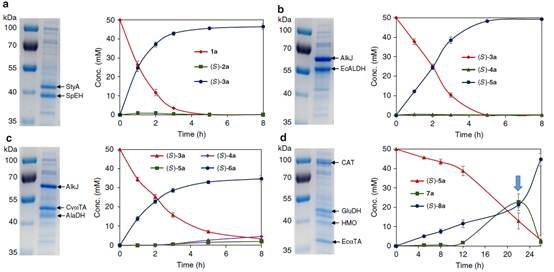
d.含有单个酶模块的大肠杆菌菌株的SDS-PAGE和生物转化时程。
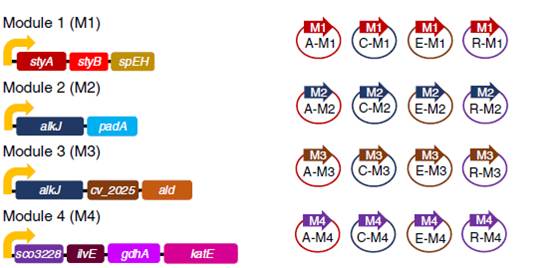
e.含有不同酶模块的重组大肠杆菌菌株的遗传构建
结论:由多个基本模块组成的有效的级联催化体系是通过不同质粒上的基本模块的组合组装开发的,以调节酶的表达水平。这些关键酶的良好的TTN值表明它们在级联生物转化中的催化效率很高。工程化的全细胞生物催化剂接受一组苯乙烯衍生物作为三种类型的不对称烯烃官能化的底物。
报告人:鞠酒报告题目:The Contribution of Non-catalytic Carbohydrate Binding Modules to the Activity of Lytic Polysaccharide Monooxygenases
报告内容:自然界中存在大量的微生物能分泌降解纤维素的酶,一些侵染植物的病原菌能分泌一些破坏植物细胞壁的糖蛋白,这些酶或蛋白通常含有非催化结构域CBM,CBMs在CAZY数据库根据其序列的特异性被分类到相应的家庭,并基于其结合底物的配体特征,分为A,B,C三种类型。这些CBM首最初在纤维素酶中被识别划分出来,它召集同源酶与相对应的底物紧密靠近,从而促进催化。这些CBMs也可以直接引导酶识别植物细胞壁上脆弱易被侵染的部位。这些结果都表明CBMs能特异性地调节酶的生物学活性。前人已经详细探讨了CBMs在糖苷水解酶中具体的功能作用。虽然30%的LPMO中存在CBM,但是CBM在LPMO中的功能相关信息很少。本文以Cellulomonas fimi和Thermobispora bispora的LPMO10为实验对象,研究了CBM对LPMO催化活性的影响。
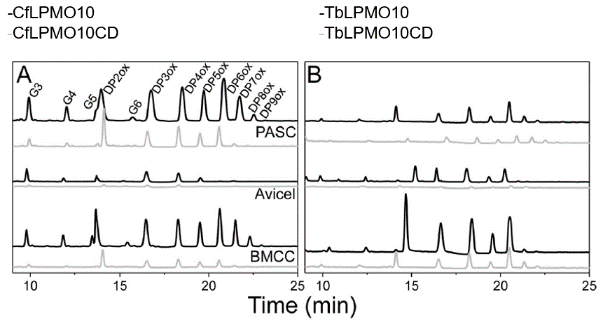
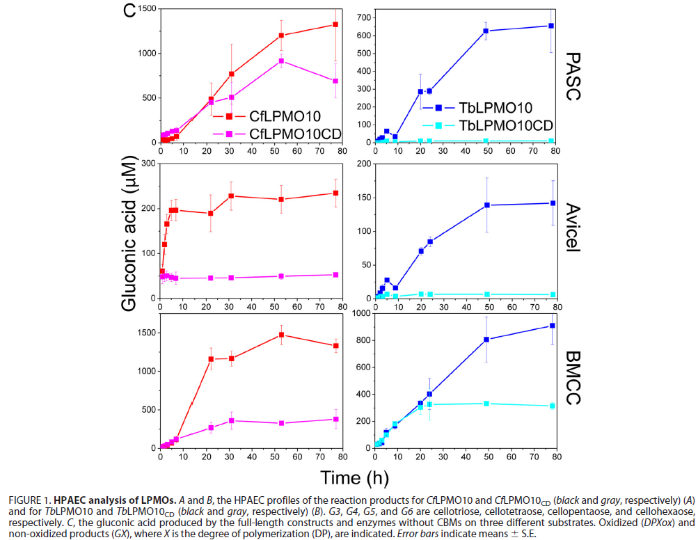
作者以Cellulomonas fimi和Thermobispora bispora的LPMO10为实验对象,首先对两个LPMO的氧化活性进行确定,如图1和图2,两种LPMO对高度结晶(Avicel和BMCC)和无序(PASC)形式的纤维素都具有活性。两种LPMO在底物为BMCC和PASC情况下比Avicel产生了更多的寡糖,这可能反映了底物表面的差异。与Avicel相比,BMCC的微纤维非常纤细薄弱, PASC前处理中酸破坏了多糖的结晶结构,增加PASC暴露在溶剂中的纤维寡糖链的数量。其次,去掉CBM的两种LPMO的截短体在以三种纤维素为底物时,寡糖产量的丰度和产量都不如天然的LPMO,说明CBM的存在对全酶的酶活性影响是积极的。
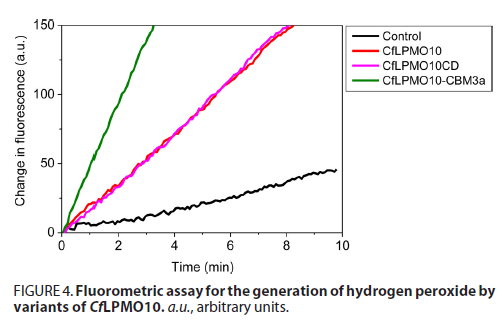
LPMO通常由催化域和非催化域通过linker连接组成全酶,传统的观点认为催化域和非催化域的功能在某种程度上是相互独立的。为了探索CBM对LPMO催化域活性的影响,作者采用了过氧化氢测定法具体测定天然LPMO和截短LPMO的活力区别,发现CBM的缺失对催化域的活性功能没有影响,这两个结构域是相互独立的。
降解纤维素和植物细胞壁的其他成分的糖苷水解酶的活性也受到的CBMs的影响。由CBM介导的活性影响似乎简单地反映了它们与底物的亲和力和结合能力。为了探究这些原理是否也适用于LPMO,LPMO与各种异源纤维素的特异性CBM相杂接,并评估所得酶的活性。下图显示,CBM对LPMO活性的影响是酶和底物特异性的。例如,在Tb LPMO10中用Cf CBM2a替代Tb CBM2a 导致对Tb LPMO10对PASC和BMCC的活性显着降低。与此相反,用CjCBM10取代Cf LPMO10的CfCBM2a,增强了对微晶纤维素的活性,但显著降低了Cf CBM2a和Tb LPMO10 对BMCC的催化能力。
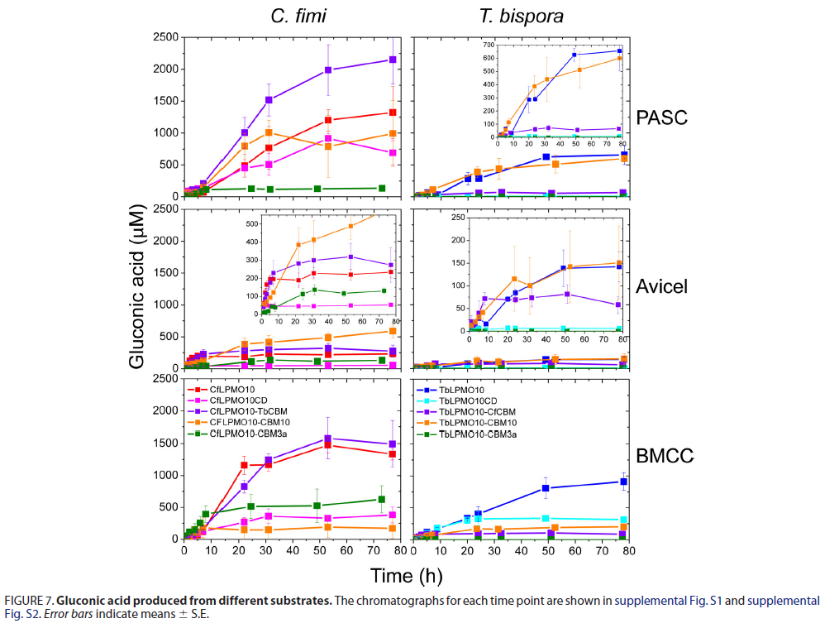
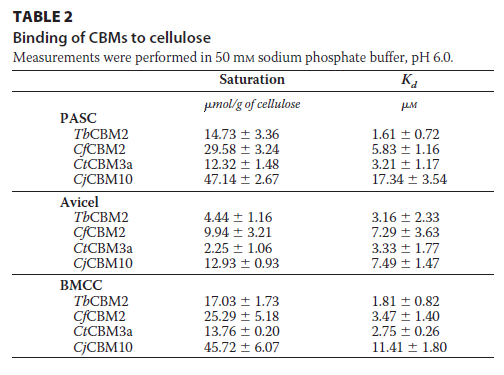
为了探索CBM的结合特性与它们对加氧酶的活力影响是否存在一定的关系,我们测定了它们的亲和和结合能力。下表和图中中提供的数据表明,截短体LPMO失去了与底物结合能力,并且外源CBM的导入能增加使LPMO恢复结合底物的能力,联系上图,结合能力与酶活力之间不存在绝对的正相关或负相关关系,而嗜热生物体CBM3a和Tb CBM2a 的两种CBM 比嗜中温细菌的蛋白质结合纤维素更紧密。可以看出,CBMs对LPMO的结合能力影响并没有为对LPMO活力的影响提供明确的见解。
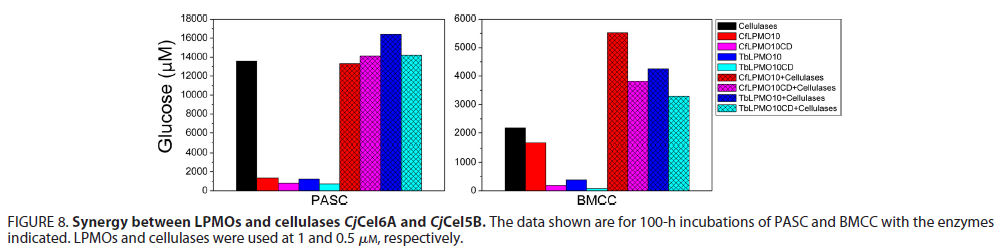
以往的研究表明,许多LPMOs能增强纤维素酶和几丁质酶对它们各自的不溶性底物的活性。对CBMs在LPMO和聚糖酶之间的协同相互作用中的作用了解甚少。为了探索这个问题,研究了杂接了不同CBMs 的与纤维二糖水解酶(Cj Cel6A)和内切葡聚糖酶(Cj Cel5B)的协同作用的能力。下图所示的数据, 表明当与纤维素酶和Cf LPMO10协同时,与将这些酶与该晶体形式的纤维素单独温育时相比,由BMCC释放的葡萄糖终产量比起单独作用时的产量显着增加。当使用PASC作为底物时,糖苷水解酶和加氧酶之间的协同作用不明显。尽管缺少CBMs的Cf LPMO10CD与纤维素酶之间的协同作用在以BMCC为底物时效果是显而易见的,所产生的葡萄糖总量降低。这些数据表明LPMO与纤维素酶的协同作用,但是这决定性的协同能力并不是由LPMO中的CBMs决定的。
总结:CBMs对LPMO的影响是依赖于LPMO特异性和底物特异性的,对C4和C1氧化产物比例的影响似乎是通过LPMO的作用模式来调节的。CBM杂接交换表明,CBMs在LPMO中扮演了更为重要的作用,不合适的CBM对LPMO活性功能甚至是消极的。纤维素底物结合研究表明,CBM的纤维素结合能力可以对LPMO活性有显着影响,并且它们的作用随着这些模块的来源,所使用的酶和所评估的底物而变化。LPMO与底物结合后在底物表面的延伸保留是由其CBM介导的。CBMs增强LPMO活性的机制比简单促进酶底物接近程度更复杂,CBM和LPMO之间的相互作用的研究是进一步探索设计高效定向组合酶时一个重要方向。

